CÁC HOẠT ĐỘNG THÍCH NGHI MIỄN DỊCH NHẰM BẢO VỆ KHỎI CÁC TÁC NHÂN GÂY BỆNH TRÊN GIA CẦM DIỄN NHƯ THẾ NÀO?
Gia cầm luôn bị đe dọa bởi các tác nhân gây bệnh như virus, vi khuẩn, động vật nguyên sinh, gian sán, ngoại kí sinh trùng (hình 1).
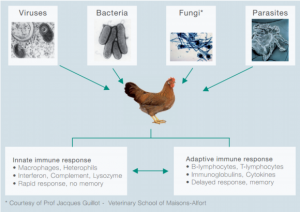
| Hình 1: Các loại tác nhân gây bệnh và hệ thống miễn dịch trên gia cầm |
Các vi sinh vật này đã phát triển riêng biệt và xâm nhiễm vào, nhân lên trong cơ thể vật chủ bằng nhiều phương thức khác nhau ,và chúng thường qua mặt được hệ thống miễn dịch.
Mặt khác, hệ thống miễn dịch đã tiến hóa như một “cơ quan” phức hợp với nhiều tầng bảo vệ nhằm phản ứng đặc hiệu với số lượng lớn các tác nhân gây bệnh.
Nó bao gồm một hệ thống phản ứng nhanh và một hệ thống phản ứng trễ và được gọi là hệ thống miễn dịch tự nhiên và hệ thống miễn dịch thu được.
Thông thường hệ thống miễn dịch tự nhiên là đủ để kiểm soát sự nhiễm khuẩn nhưng nếu nó không đặc hiệu, thì miễn dịch thu được sẽ được kích hoạt. Cần lưu ý rằng cả hai hệ thống đều tương tác với nhau chặc chẻ; qua các ví dụ sẽ trình bày sau đây.
Hệ thống miễn dịch thu được bao gồm tế bào lympho B và T với khả năng đặc biệt là có thể ghi nhớ được các mầm bệnh trước đó và có thể nhận diện lại các mầm bệnh khi bị tái nhiễm, do đó hệ thống miễn dịch thu được được cho là nhanh và có hiệu quả cao.
Bài viết này tập trung vào hệ thống tế bào lympho B. Đầu tiên là tóm lược cấu trúc và chức năng của kháng thể, sản phẩm bài tiết của tế bào B. Kế tiếp là sự phát triển của tế bào B trong quá trình phát triển của phôi và thảo luận về vai trò của túi bursa Fabricius.
Cuối cùng, giải thích về sự kích hoạt tế bào B nhằm tạo ra kháng thể và khả năng ghi nhớ của hệ miễn dịch. Vai trò của tế bào lympho T sẽ chỉ được đề cập đến trong vai trò hỗ trợ cho tế bào B.
Kháng thể
Kháng thể có bản chất là protein và được tiết ra bởi tế bào B và tương bào trong mô và máu. Bởi vì nó có cấu trúc hình cầu nên cũng còn được gọi là immunoglobulins.
Trên gà có ba loại immunoglobulins đó là: IgM, IgY và IgA (hình 2). Các loại chim thiếu sự tương đồng với động vật có vú vì động vật có vú có thêm phân tử IgE.
Cấu trúc đơn giản của tất cả immunoblobulins là hình thái Y và bao gồm hai chuỗi nhẹ (light chain) và hai chuỗi nặng (heavy chain) được liên kết với nhau bởi cầu nối disulfide (hình 2A).
Trên một đầu của phân tử immunoglobulin có thể gắn với các kháng nguyên trong khi đầu còn lại có chức năng phản ứng trung hòa kháng nguyên khi được gắn kết.
Immunoglobulin đầu tiên được sản xuất trong quá trình phản ứng miễn dịch thì được gọi là IgM (hình 2B). Trong phân tử này có năm hình thái Y được liên kết với nhau tạo thành một phân tử năm nhánh (pentamer) và cho phép phân tử có thể gắn kết với nhiều kháng nguyên cùng một thời điểm.
IgM phần lớn được tìm thấy trong máu và một phần nhỏ trong protein được tiết ra trên bề mặt niêm mạc.
Trong quá trình nhiễm khuẩn lần đầu và phản ứng của nhiễm khuẩn thứ phát, IgY trở thành một immunoglobulin nổi bật nhất. IgY là một phân tử một nhánh (monomer) và được tiết ra với nồng độ cao bên trong máu (7-12 mg/ml), nhưng nó được tìm thấy một ít trên bề mặt niêm mạc.
Trong quá trình hình thành lòng đỏ trứng, IgY được truyền từ máu của gà mẹ sang lòng đỏ trứng. Phôi phát triển hấp thụ dưỡng chất từ lòng đỏ đồng thời IgY được truyền vào hệ tuần hoàn của phôi nhằm bảo vệ gà con trong thời điểm quan trọng đầu tiên sau khi nở.
Trái ngược với IgM và IgY, IgA được tiềm thấy với số lượng ít trong máu. Tuy nhiên, IgA là immunoglobulin nhiều nhất trong cơ thể từ khi nó được tế bào lympho B ở bề mặt niêm mạc tiết ra với số lượng nhiều và được bài tiết trực tiếp như là một phân tử hai nhánh (dimer) hoặc phân tử ba nhánh (trimer) vào bề mặt niêm mạc để ngăn chặn các vi sinh vật xâm nhập vào.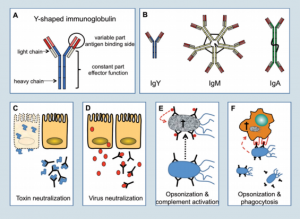
| Hình 2: Các loại kháng thể và cơ thế hoạt động |
Kháng thể hoạt động theo nhiều cách khác nhau để kiểm soát sự nhiễm trùng. Tuy nhiên, đa số những hoạt động của kháng thể giới hạn ở không gian ngoại bào. Các tác nhân gây bệnh nội bào không được kiểm soát tốt bằng kháng thể; ví dụ như những loại virus nhân bản bên trong tế bào và vi khuẩn nội bào như Salmonella, Listeria hoặc Mycobacteria.
Tuy nhiên đối với môi trường ngoại bào và trên bề mặt niêm mạc, kháng thể có thể gắn kết vững chắc với các phân tử hòa tan hoặc thành phần cấu trúc của các vi sinh vật.
Sự gắn kết của kháng thể với vi khuẩn hoặc độc tố nấm có thể trung hòa được các độc tố và được biết đến từ kháng thể chống độc tố uốn ván trong quá trình tiêm chủng (Hình 2C).
Trong trường hợp của virus, kháng thể có thể gắn kết với kháng nguyên của virus trong quá trình nhiễm trùng máu (IgY) hoặc trên bề mặt niêm mạc (IgA) và ngăn chặn lây nhiễm của các tế bào trong việc ức chế sự tương tác giữa virus với tế bào (Hình 2D).
Ví dụ kháng thể trực tiếp chống lại hemagglutinin có trong virus cúm đã hoạt động theo cơ chế trên. Sự gắn kết của kháng thể với vi khuẩn có thể diệt được vi khuẩn bằng nhiều cách khác nhau.
Lớp bọc của vi khuẩn với kháng thể được gọi là opsonisation ( opsonisation: là lớp bọc opsonin, một thành phần trong huyết thanh, nhằm làm cho tế bào vi khuẩn hấp dẫn các Thực bào) và chúng kích hoạt các Bổ thể và cuối cùng vi khuẩn bị phân giải bởi những thành phần bổ thể đó (Hình 2E). Ngoài ra, Lớp bọc vi khuẩn được nhận diện bởi bạch cầu trung tính, đại thực bào những yếu tố sẽ diệt được những mầm bệnh.
Để xác định được vi sinh vật opsonised, những thực bào sẽ thực hiện trên các thụ thể bề mặt nơi mà gắn kết với các kháng thể phủ bề mặt của mầm bệnh (Hình 2F). Không một cơ chế nào có thể thay thế được trên động vật bị thiếu hụt kháng thể. Vì thế sự thiếu hụt kháng thể sẽ gây ra hậu quả nghiêm trọng đe dọa đến mạng sống.
Tế bào lympho B phát triển ở túi Bursa
Hệ thống tế bào B phát triển trong quá trình hình thành phôi và một vài tuần đầu sau khi nở. Cơ quan trung tâm của quá trình này là túi Bursa Fabricius và được nối bởi ống mở đến hậu môn (Hình 3)
Nó cung cấp môi trường cho tế bào B chưa trưởng thành phát triển thành các tế bào lympho B có chức năng sản xuất các kháng thể đặc hiệu khác nhau.
Do đó, khi phẩu thuật cắt bỏ túi bursa hoặc phá hủy bằng hóa chất, chất độc hoặc virus có thể dẫn đến tình trạng suy giảm miễn dịch nghiêm trọng và làm tăng tính nhạy cảm với các tác nhân gây bệnh trên gia cầm.
Trong thời điểm 8-15 ngày tuổi trong quá trình hình thành phôi (ED), tiền tế bào B bursa (tế bào B gốc) di chuyển từ lá lách của phôi thai và tủy đến bên trong túi bursa. (Hình 3.1).
Ở đây nó nhận được tín hiệu từ các mô bursa nơi mà kích hoạt quá trình trưởng thành. Bản chất của những tín hiệu này vẫn còn chưa được hiểu rỏ. Tuy nhiên, Những hoạt động xảy ra trong quá trình phát triển tế bào B để phản ứng với các tín hiệu này đã được nghiên cứu sâu rộng.
Ban đầu túi bursa được hình thành bởi vài ngàn tiền bursa như một bước đầu tiên, sau đó bắt đầu phát triển và hình thành khối tế bào B được gọi là nang bursal (Hình 3.2). Ước tình rằng một bursa trưởng thành có đến 12000 nang với khoảng 2×105 tế bào B trên một nang.
Trong quá trình tăng sinh của tế bào B, một quá trình quan trọng thứ hai được bắt đầu dẫn đến sự hình thành của hàng tỉ tế bào lympho B, mỗi tế bào sản xuất ra một phân tử kháng thể đặc hiệu với khả năng gắn kết đặc hiệu.
Khi tế bào đã hoàn toàn trưởng thành, thế bào B sẽ rời khỏi cơ quan bursa và đến mô ngoại biên. Quá trình này bắt đầu từ lúc gia cầm nở và được tiếp diễn đến khi túi bursa biến mất trong thời điểm thành thục của gia cầm.
Tuy nhiên, sự phát triển của túi bursa chưa được hoàn thiện tại quá trình nở. Trong hai tuần đầu sau khi nở, nang được thay đổi cấu trúc với sự phát triển của vỏ nang và tủy (Hình 3.3).
Người ta cho rằng chức năng của tủy như là một cơ quan bạch cầu thứ cấp rất giống với lá lách trong khi vỏ sản xuất ra tế bào B mới và tiếp tục lưu trú ở cơ quan bạch cầu ngoại biên (Hình 3.4).
Phẩu thuật loại bỏ bursa chỉ ra rằng đây là cơ quan cực kì quan trọng đến khi gà đạt đến độ tuổi ít nhất là 4-5 tuần.
Vào thời điểm đó số lượng tế bào B bắt đầu xuất hiện bên trong lá lách có thể duy trì các hệ thống tế bào B của gà khi túi bursa đã được gỡ bỏ. Tuy nhiên, bursa vẫn còn cần thiết trong những tuần sau đó để đảm bảo một hệ thống tế bào B đầy đủ chức năng.
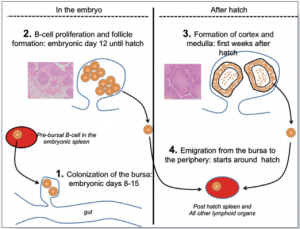
| Hình 3: Vai trò của bursa |
Chức năng của tế bào B trong cơ quan bạch cầu thứ cấp.
Tế bào B rời khỏi bursa và di chuyển đến cơ quan bạch cầu thứ cấp, đặc biệt là lá lách, hạch manh tràng (Caecal tonsils) ở ruột, phế quản có liên quan đến mô bạch huyết ở phổi (BALT) và hạch bạch huyết ở trong mô niêm mạc.
Tại đây những tế bào tương tác với tác nhân gây bệnh hoặc kháng nguyên được đưa vào bằng đường tiêm chủng. Như đã nêu trước đây, tế bào B phát triển trong bursa cho ra hàng tỉ tế bào sản xuất kháng thể có khả năng gắn kết những kháng nguyên khác nhau.
Phân tử immunoglobulin này được giữ ở màng tế bào với vị trí gắn kháng nguyên đối diện với môi trường (Hình 4, phần trên).
Nó là một immunoglobulin loại IgM được thể hiện như một monomer và được xem là thụ thể kháng nguyên tế bào B hoặc BCR. Sự gắn kết của kháng nguyên thích hợp với BCR sẽ kích hoạt tế bào B, thúc đẩy sự tăng sinh và sự khác biệt với kháng thể tiết ra tế bào tương.
Trong quá trình này, tế bào tương sẽ mất BCR trên bề mặt tế bào nhưng bắt đầu tổng hợp một lượng lớn phân tử immunoglobulin và tiết ra bên trong mô và máu. Quá trình kích hoạt này cần đến 5-6 ngày, nó giải thích tại sao sự xuất hiện kháng thề IgM đặc hiệu kháng nguyên đầu tiên được nhận biết gần 1 tuần sau khi tiêm chủng.
Một đặc điểm đặc trưng của kháng thể IgM là ái lực kháng nguyên thấp đó là kết quả từ một sự phù hợp không hoàn hảo giữa kháng nguyên và sự gắn kết của phân tử IgM. Nhược điểm này thay thế bằng cấu tạo pentameric của sự bài tiết IgM và cho phép đồng thời gắn kết nhiều vị trí do đó làm tăng sự gắn kết chung.
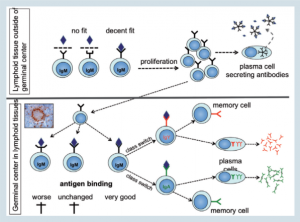
| Hình 4: Thích ứng |
Hai đặc điểm của phản ứng kháng thể đã thu hút các nhà miễn dịch học trong một quãng thời gian dài. Đầu tiên là sự xuất hiện cùa kháng thể IgY hoặc IgA trong quá trình phản ứng với vị nhiễm khuẩn, quá trình này được gọi là “chuyển đổi lớp immunoglobulin”
Thứ hai là sự sản xuất của kháng thể với liên kết kháng nguyên mạnh hơn nó được gọi là “affinity maturation”(thuần thục ái lực). Cả hai tiến trình đều xảy ra trong một cấu trúc ở mô bạch huyết (the germinal center – GC).
Ở chim nó là những cấu trúc vòng được tìm thấy ở lá lách, hạch manh tràng và BALT (Hình 4). Sau cuộc gặp đầu tiên với kháng nguyên một vài tế bào tăng sinh sẽ bắt đầu hình thành GCs.
Trong quá trình này các tế bào sẽ tiếp tục phân chia và cùng một thời điểm sẽ bắt đầu biến đổi immunoglobulin gen (Hình 4; phần bên dưới).
Những đột biến này sẽ diễn ra tại vị trí gắn kháng nguyên và thay đổi nó theo hướng mà sức mạnh gắn kết có thể không đổi hoặc thấp hơn hoặc cải thiện tốt hơn.
Chỉ có những tế bào B với gen immunoglobulins cải thiện sẽ tiếp tục tăng sinh và trong bước tiếp theo sẽ nhận tín hiệu để thay đồi từ IgM sang IgY hoặc IgA (chuyển lớp). Cuối cùng những tế bào này sẽ phân biệt với tế bào tương và bắt đầu tiết ra một lượng lớn immunoglobulin.
Kết quả cuối cùng của quá trình này là thế hệ của kháng thể mà có thể “bắt lấy” kháng nguyên mạnh hơn và do đó có thể loại bỏ được các tác nhân gây bệnh hiệu quả nhất.
Đây là một cơ chế quan trọng minh họa bằng sự tăng cao độ nhạy cảm với nhiễm khuẩn ở bệnh nhân với việc thất bại trong quá trình chuyển lớp. Mặc dù hiếm nhưng nó vẫn được mô tả ở ngưởi và chuột mà còn ở loài gia cầm.
Nó cần phải được lưu ý rằng quy trình này không được thể tiến hành tế khi vắng mặt tế bào T. Một tế bào Tín hiệu quan trọng của “thuần thục ái lực (affinity maturation)” và “chuyển lớp (class switching)” được cung cấp bới tế bào T-helper.
Do đó, sự vắng mặt của các tế bào T-helper, hoặc suy chức năng, sẽ gây suy giảm miễn dịch nghiêm trọng trong hệ thống tế bào B.
Trong khi thành thục ái lực và chuyển mạch lớp giúp cải thiện bảo vệ, sự hình thành của các tế bào bộ nhớ sẽ giúp động vật để đáp ứng rất nhanh với sự tiết ra ái lực cao kháng thể IgY hoặc IgA trong quá trình tái nhiễm.
Tế bào bộ nhớ được tạo ra trong GCs trong quá trình nhiễm khuẩn lần đầu (Hình 4) và có thể sống trong vài tháng đến vài năm.
Ở chuột và người những tế bào này tồn tại ở tủy. Vị trí của nó ở gia cầm vẫn chưa được rõ nhưng sự tồn tại của nó là chắc chắn có. Tế bào bộ nhớ là mục đích đầu tiên trong việc tiêm chủng.
Việc tiêm chuẩn đầu sẽ dẫn đến việc hình thành các tế bào bộ nhớ mà được kích hoạt lại trong quá trình tái tiêm chủng hoặc trong trường hợp của nhiễm khuẩn bởi cùng một loại tác nhân gây bệnh trước đó.
Phản ứng thứ phát này tạo ra một lượng lớn tế bào B có nguồn gốc từ tế bào bộ nhớ B mà trong đó việc sản xuất một lượng lớn kháng nguyên với khả năng gắn kết mạnh. Ngoài ra, các tế bào bộ nhớ mới được hình thành, để sẵn sàng cho một thách thức tiếp theo của những mầm bệnh tương ứng.
References
- Härtle S, Magor K.E., Göbel, T.W., Davison F. Kaspers B. Structure and evolution of avian immunoglobulins. pp 103- 120; In: Avian Immunology, Schat K.A., Kaspers B., Kaiser P. 2nd edition, Elsevier 2013.
- Oláh I, Nagy N. Retrospection to discovery of bursal function and recognition of avian dendritic cells; past and present. Dev Comp Immunol. 2013, 41(3):310-5
- Ratcliffe M.J.H., Härtle, S. B cells, the bursa of Fabricius and the generation of antibody repertoires. pp 65- 90; In: Avian Immunology, Schat K.A., Kaspers B., Kaiser P. 2nd edition, Elsevier 2013.
- Toivanen P, Toivanen A. Bursal and postbursal stem cells in chicken. Functional characteristics. Eur J Immunol. 1973, 3(9):585-95)
Biên soạn: Phòng chẩn đoán và phòng ngừa dịch bệnh – Công ty cổ phần 3F VIỆT.
Source: